الأدلة المؤيدة للتطور: قراءة في السجل الأحفوري والوراثة (2/6)

أحد أقوى أسباب قبول كثير من العلماء لنظرية التطور هو اتحاد أدلة مستقلة ومتنوعة كلها تشير إلى نفس الاستنتاج: أن الكائنات الحية تتغير عبر الزمن، وأن بعض هذه التغيرات تُورَّث وتنتج أنواعًا جديدة عبر عمليات مثل الطفرات والانتقاء والانتشار الجيني. في هذا المقال نفصل هذه الخطوط الدليلية، ونقدم أمثلة ملموسة مدعومة بمراجع علمية.
1 — الأدلة الحفريّة: أمثلة انتقالية تُظهر تغيّر الأشكال عبر الزمن
السجل الأحفوري لا يعطي سلسلةً كاملةً لكل نوع، لكنه يقدم ما يعتقد أنها أمثلة انتقالية تُظهر صفات وسيطة بين مجموعات كبرى:
- Tiktaalik roseae: اكتشاف اكتمل وصفه في 2006 أظهر هيئة سمكية ذات وهج من خصائص الأطراف الأولى (مفاصل قريبة للمعصم/الكوع) وميزات للزحف على الماء الضحل — جسر واضح بين الأسماك والرباعيات البرية. (راجع المقال الأول من السلسلة – فقرة 7)
- Archaeopteryx: أحفورة شهيرة تجمع بين سمات الديناصورات (أسنان، ذيل عظمي) وسمات الطيور (ريش، عظمة قص) — تُستخدم عادة كمثال تقليدي على شكل انتقالي بين الديناصورات والطيور. (راجع المقال الأول من السلسلة – فقرة 7)
- التحول من ثدييات برية إلى حيتان: اكتشافات مثل Pakicetus، Ambulocetus، وBasilosaurus تظهر سلسلة انتقالية من ثدييات أرضية شبه مائية إلى حيتان بحرية متكيفة تمامًا — تفاصيل تشريحية توافق توقعات نسبٍ مشتركة.
- سجل الخيول: تطور الخيول يقدم تسلسلاً أحفوريًا غنيًّا من أحجام أصغر وأصابع متعددة إلى الخيل الحديث مع إصبع مفرد وتكيفات للرعي والجري. هذا السجل يُعد من أكثر سجلات الحفريات اكتمالًا ويُستخدم كمثال تعليمي لصيغة التغيرات التطورية عبر الزمن.









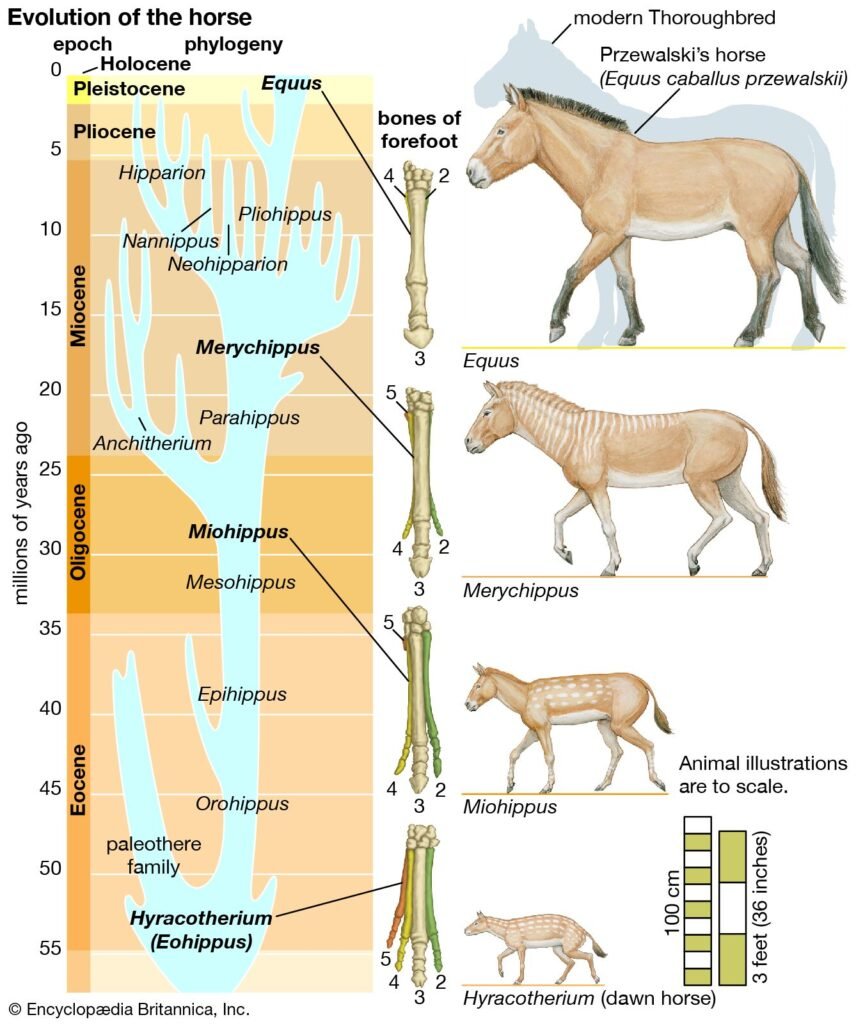
الاستنتاج: الحفريات تُظهر أشكالاً انتقالية متعددة على مستوى مجموعات كبيرة، وهذه الأمثلة تدعم فكرة النسب المشترك والتغيير التدريجي/التبقّي-الطبيعي.
2 — التشابه البنيوي (تشريحيّاً) والتعاقب الجنيني (Embryology)
- الهومولوجيا (Homology): هياكل تشريحية متشابهة في الكائنات المختلفة (مثل عظام طرف الإنسان والحصان والحوت والطيور) تفسرها الفرضية القائلة بوجود أصل مشترك ثم تباين وظيفي لاحق.
- وظيفة التطابق مقابل التقارب (Convergence): لا بد من التفريق بين أوجه التشابه الناتجة عن أصل مشترك (homology) والتشابه الناتج عن حلٍ شكلّي مستقل استجابةً لضغوط بيئية مماثلة (convergent evolution) — كلا الحالتين موثقة ومعروفة.
- علم الأجنة: تشابه مراحل الجنين بين الفقاريات في مراحل مبكرة (مثل وجود خياشيم/شقوق بلعوميّة/ذيل خلال تطور الجنين) يشير إلى قواعد تنظيمية مشتركة في التطور والتطور الجنيني. (لاحظ أن التفسيرات الحديثة تُركِّز على آليات جزيئية مشتركة تتحكم في هذه الأنماط).
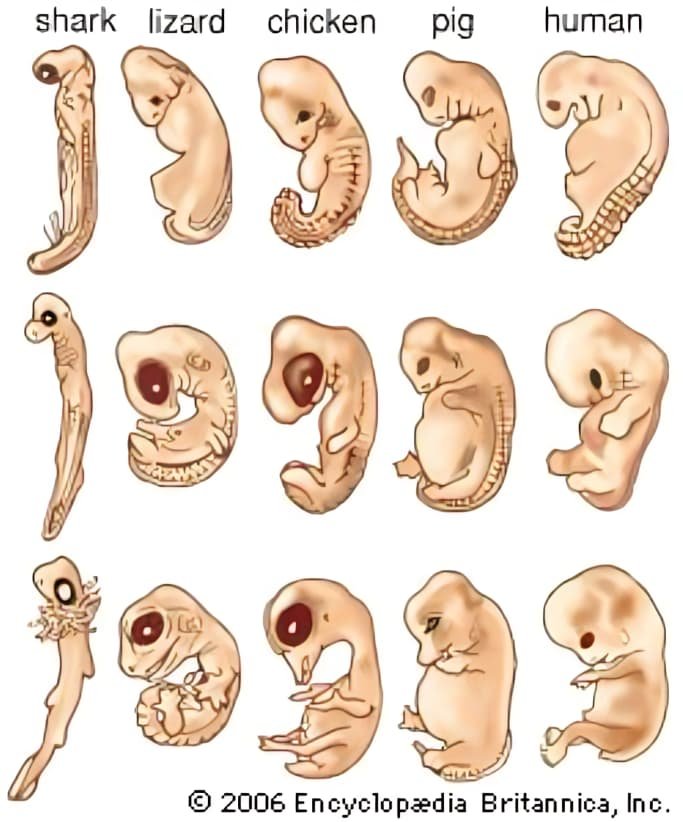
3 — الأدلة الجزيئية والجينية (الـDNA وبيانات الجينوم)
في العقود الأخيرة، قدّم علم الجينوم أدلة قوية تدعم النسب المشترك:
- التشابه الجيني العام: مقارنة تسلسلات DNA و البروتينات بين الأنواع تُظهر أن الأنواع الأقرب في الشجرة التطورية تتشارك في نسب أعلى من التسلسلات؛ هذا يتوقعه نموذج النَسْب المشترك. (مقارنة الإنسان/الشمبانزي مثالٌ بارز في هذا السياق).
- العناصر الفيروسية الذاتية (Endogenous Retroviruses — ERVs): أجزاء من فيروسٍ اندمجت في جينوم أسلافنا وصارت موروثة — وجود نفس العناصر في مواقع متطابقة في جينوم الإنسان والشمبانزي يُفسَّر بأن هذه الإدخالات حدثت في سلف مشترك ثم وُرِّثت. هذا النوع من الدليل يُعتبر من أقوى الشواهد الجزيئية على أصلٍ مشترك.
- المُزوّفات (pseudogenes): جينات تعطّلت (أصبحَت غير فاعلة) في نفس المواضع في جينومات أنواع متقاربة — مشاركة نفس الطفرات المعطِّلة تُشير إلى أصل مشترك. هناك نقاشات متخصصة حول وظائف بعض pseudogenes، لكن نمط التوزيع الداعم للأصل المشترك واضح في كثير من الدراسات الجينومية.
- تكرار الجينات والابتكار الجيني: آليات مثل تكرار الجينات (gene duplication) ثم تمايز النسخ أعطت موادًّا خامًا لظهور وظائف جديدة — فكرة مركزية منذ كتاب Susumu Ohno (1970) وما تلاها من دراسات جينومية.
النتيجة: الأدلة الجزيئية تعمل كخط مستقل يتقاطع مع الحفريات والتشريح (consilience)، مما يجعل الاستنتاج التطوري أكثر إحكامًا.
4 — أمثلة رصدية وتجارب تُظهر التطور في الزمن الحقيقي
النظرية ليست فقط تفسيرًا لسجلٍ تاريخي — هناك حالات رُصدت مباشرة أو في تجارب مخبرية:
- تجربة E. coli طويلة الأمد (Lenski LTEE): تجربة بدأت على نطاق أجيال عديدة أظهرت تغيّرًا وراثيًا وتكيّفات مفيدة، ومنها ظهور قدرة متمَيِّزة على استخدام السيتريت بصورة هوائية في إحدى السلالات (ظاهرة نُشرت وناقشت بتفصيل في تقارير الفريق). هذه التجربة تُظهر كيف تتراكم الطفرات وتنتج وظائف جديدة تحت ضغوط انتقائية.
- طيور داروين (دراسات Grants): أبحاث طويلة الأمد على جزر غالاباغوس مرّت بتوثيق تغيرات سريعة نسبياً في أحجام وبعض سمات المنقار استجابةً لتغير موارد الغذاء والضغوط البيئية، وأظهرت أيضًا آليات تساهم في العزل الجنسي/التكوّن النوعي. هذه مشاهدة ميدانية قوية لآليات الانتقاء الطبيعي. (راجع المقال الأول من السلسلة)
- المقاومة للمضادات الحيوية والتكيفات الجرثومية: تطور مقاومة بكتيرية (مثل MRSA وأنماط مقاومة أخرى) عبر طفرات وانتقال جينات مقاومة — مثال عملي وواضح على كيف تغير الضغوط البيئية ترددات الجينات لصالح بدائل قادرة على النجاة. هذا التطور يوضح أيضًا دورُ التجمعات السريعة و/أو الانتقال الأفقي للجينات في تغيير المشهد التطوري.
- حالة العثة (Biston betularia) — melanism: قصة العثة السوداء في بريطانيا أثناء الثورة الصناعية كانت مثالاً تعليميًا مشهورًا على الانتقاء الطبيعي (تغيّر توزع الألوان ضد خلفية ملوّثة)، مع مناقشات منهجية وتفاصيل لاحقة حول طرق الاختبار الميداني، ولكنها لا تزال مثالاً مهمًا لتأثير البيئة على ترددات الطفرات المفيدة.
الخلاصة: دراسات الميدان والتجارب المخبرية تُظهر عملياتٍ تطورية قابلة للقياس ضمن أزمنة قصيرـة إلى متوسطة، وتعمل كحجج مباشرة تدعم آليات التطور.
5 — علم الأنماط التطورية والجُذور التطورية (Phylogenetics & Molecular Clocks)
علم الأنساب الجزيئية يستخدم تسلسلات جينية لبناء شجرات تطورية تُقدّر العلاقات الزمنية بين الأنواع:
- طرق تحليلية متقدمة: تقنيات إحصائية مثل التحليل بأقصى احتمال (maximum likelihood) أو الأساليب البايزية تُستخدم لبناء أشجار موثوقة من تسلسلات DNA/البروتين، مع برامج متطورة (مثلاً MrBayes). هذه الأساليب تقلل الأخطاء الناتجة عن اختلاف معدلات التطور وتسهم في اختبار فرضيات النسب
- الساعة الجزيئية (Molecular clock): فكرة مفادها أن بعض الجينات تتراكم فيها تغيّرات تقريبًا بوتيرة مستقرة في فترات زمنية طويلة؛ تُستخدم هذه الفكرة (مع تقويم بالبيانات الأحفورية) لتقدير توقيت الفواصل التطورية. هناك تحسينات وقيود (معدلات مختلفة بين الجينات والسِلالات)، لكن الأسلوب أثبت فائدته في العديد من الفروع.
هذه الأدوات الجزيئية تُكمّل الحفريات والتشريح وتمنحنا قدرةً على اختبار فرضيات تاريخية بدقة أكبر.
6 — علم التطور التطوري-التنموي (Evo-Devo) ودور جينات التنظيم (Hox / Pax6)
حقل الـevo-devo كشف أن مجموعة صغيرة من جينات التحكم في النمو (toolkit genes) تُستخدم مرارًا وبطرقٍ مختلفة لبناء أشكال متعددة. أمثلة واضحة:
- Hox genes تلعب دورًا رئيسيًا في تنظيم المحور الجسماني للأعضاء عبر الحيوان كله، وتوضّح كيف يمكن لتغييرات في أنماط التعبير أن تنتج اختلافات شكلية كبيرة.
- PAX6 (أو ey/Pax6) هو مثال قوي: نفس الجين مرتبط بتطور العين في مجموعات حيوانية متباعدة، ما يشرح كيف أن عناصر تنظيمية جزيئية قد تُستخدم مرارًا لبناء أعضاء تشبه بعضها عبر مجموعات مختلفة — دلالة على أصل تطوري مشترك للأدوات التطورية.
Evo-Devo يشرح جزءًا من كيفية حدوث الابتكار التطوري (إعادة توظيف وحدات جينية تنظيمية بدل اختراع قطعٍ جديدة بالكامل)، وهو ينسجم مع الملاحظة الجينية والفسلجية.
7 — تجميع الأدلة (Consilience) ولماذا هذا مهمّ
أهم ما يميّز حالة الأدلة لصالح التطور هو اتساق مستقل لخطوطٍ متعددة: الحفريات، التشابه التشريحي، الأنماط الجنينية، الأدلة الجزيئية (ERVs، pseudogenes، تسلسل البروتينات)، البيانات التجريبية والميدانية، وطرق الاستدلال الجزيئي كلّها تشير إلى نفس نموذج التفرّع والنسب المشترك. هذا التلاقي يُعرف بالـconsilience ويقوّي الثقة العلمية في الاستنتاج العام.
8 — القيود والمناظرات العلمية (نقطة موضوعية)
لا يعني هذا أن كل شيء “مُحلّ” — هناك حدود وأسئلة نشطة في الأبحاث:
- سجل الحفريات لا يزال غير مكتمل في كثير من المجموعات؛ اماكن وظروف الترسيب تحدد ما يُحفظ وما يُفقد.
- معدلات التطور الجزيئي تختلف بين الجينات والسلالات، مما يطرح تحديات لساعة جزيئية دقيقة مطلقة.
- الانتقال الأفقي للجينات (horizontal gene transfer) عند الميكروبات يُعقّد بناء شجرة تفرّعية واضحة لكل الحياة.
- بعض التركيبات المعقّدة جداً (مثل شبكات تنظيمية متعددة المكونات) لا تزال تُبحث عن تفسيرات تفصيلية لكيفية تشكّلها خطوة بخطوة.
مع ذلك، وجود هذه القضايا لا يقلل من قوة الأدلة المتعددة التي تلتقي على نموذج عام للتغيير والتفرُّع البيولوجي.
الأدلة المؤيدة لنظرية التطور كثيرة ومتشعبة: من حفريات انتقالية ملموسة، إلى دلائل جينية مفصّلة (بما في ذلك عناصر فيروسية وراثية مشتركة)، إلى رصد وتوثيق تطورٍ فعلي في الحقول والمختبرات. هذه الخطوط المتعددة تعمل معًا لتعطي تفسيرًا متماسكًا لتنوع الحياة. في المقال الثالث سننتقل إلى الأدلة المعارضة للتطور: مراجعة علمية شاملة: نقاط الضعف المعتبرة، الأسئلة البحثية المفتوحة (أصل الحياة، تطور التعقيدات الجزيئية العالية، بعض تفاصيل تكوّن الأنواع الكبرى)، وكيف يردّ المجتمع العلمي على هذه التحديات.



